|
1. はじめに
遺跡出土木材などの編年を行う上で年輪解析は非常に有効な手段である。出土材などの年代関係を知る方法として、土層や切りあい、放射性炭(C14)年代による年代決定などがあるが、高い精度での年代決定は困難である。これに対し年輪を使った年輪年代では出土した木材の相対的な年代関係を年単位で明らかにすることが出来る。それにより当時の人口の推移や生活を知る上で貴重な情報を得ることが出来る。木村ほか(2004)は縄文晩期終末の青田遺跡で出土した掘立柱建物の木柱を使った年輪解析を行い、建物間の年代関係を年単位で明らかにした。また、現生から過去にわたる長期的な年輪のものさしである標準パターンと出土木材の年輪パターンを合わせることにより暦年代を決定することも可能である。日本ではスギ、コウヤマキ、ヒノキにおいて標準パターンがつくられている(光谷1987)。欧米では多くの遺跡においてこうした年輪年代学的解析が行われているが、日本では遺跡出土木材を用いた年輪解析はほとんど行われていない。そのため、長期にわたる標準変動パターンのためのデータが不足している。しかし、標準パターンが無い場合、放射性炭素(C14)年代と年輪年代を併用したC14ウイグルマッチングによって高精度での暦年代決定を行うことが可能である。
姫御前遺跡IIでは古墳時代前期の掘立柱建物、木柱列、平地式建物とスギやトネリコなどの埋没林が見つかった。建物の年代は遺物や建築様式によって埋没林は層位によって判断されたが、建物間や埋没林との詳細な年代関係は明らかになってはいない。建物と隣接した場所から埋没林が見つかっているため、建物の木材をこの埋没林から切り出した可能性もあるが、年代関係が分からないため議論することが出来ない。本研究では年輪年代学を用いた、木柱、埋没林の年代関係結果、埋没林と木柱とのC14 ウイグルマッチング法による暦年代結果を示す。
2. 試料
試料は姫御前遺跡IIより出土した古墳時代前期の建物、木柱列を構成する8本及び埋没林である自然木191点(うち一点は竹花遺跡出土)である。自然木は年輪数が少ない枝材は除く。竹花遺跡は姫御前遺跡から200mほど離れた場所に位置し、年代が重複するため断続的な一連の遺跡ではないかと考えられている。樹種同定は東北大学で行われた。
木柱はP340が掘立柱建物(SB341)、P245,P246,P247,P248,は平地式建物(SI216)、P420,P421,SX224が木柱列(SA199)を構成する柱である。
3. 方法
3−1 年輪計測法
切り出されたサンプルの切断面(木口面)を耐水サンドペーパーを付けたサンダーを用いて流水下で研磨し、年輪を読みやすくした。その後A3のフラットベッドスキャナ[エプソンES−10000G]を用いて画像を取り込んだ。解像度は1200dpiを標準として、年輪の詰まりなどの場合2400dpiとした。次に画像上で複数方向(主に3方向a,b,c)の側線をとり、年輪計測ソフト「ringmeas」を使って外側から中心へ向かって1年輪毎の幅を計測した。画像上で確認しにくい場合は双眼実体顕微鏡を用いてサンプル自体で確認を行った。
また、広葉樹(モクレン属ほか)において年輪のコントラストが低く、上記方法で確認できないものについては、薄い切片を作成して透過光を読み取る方法を用いた。この場合サンプルの最外部から中心へ1〜2cmほどの木口面に対して水平になるよう出来るだけ薄い切片をとり、中心へ向かって複数切片をそれぞれのつながりが分かるように採取した(図1)。その後、透過原稿ユニットを内蔵したスキャナ[エプソンGT-9800]で画像を解像度3200dpiで取り込み、年輪計測ソフト「ringmeas」を使って外側から中心へ向かって1年輪毎の幅を計測した。計測後、個体内でとった複数の側線データをクロスデーティングソフト[cross9](Kimura 1995)を用いて読み間違いが無いか検証後側線データを平均化、その個体の年輪データとした。この際、個体内での相関が悪い個体は、偽年輪や不連続年輪、読み間違え等が無いか画像上で年輪界をたどるなどして確認した。この確認後も偽年輪等により、個体内の同調性に信頼性が無い個体についてはその後の年輪解析からは除いた。
3−2 試料間の相対年代関係の決定
木柱や自然木の個体間の相対的な年代関係を調べるため、平均化した個体データを元に個体間をクロスデーティングソフト[cross9] (Kimura 1995)でクロスデーテトし、その相関を調べた。標準化処理には主に5年移動平均法を用い、クロスデーティングの相関の有意性を示すt値(Baillie 1982)は5を目安とした。自然木に関しては同一樹種の個体間、木柱に関しては同一建物木柱間、異なる建物木柱間でクロスデーティングを行い、相対的な年代関係を決定した。その後、年輪パターンを平均化し、マスタークロノロジーをそれぞれ作成した。
3−3 ウイグルマッチングによる暦年代決定
年輪年代では出土材の相対的な年代関係は求められるが、暦年代決定にはものさしとなる標準パターンが必要である。しかし、この標準パターンが無いため年輪年代学で暦年代決定を行うことが出来ない。そこでC14年代測定とC14年代較正曲線(Intcal04)を利用したC14ウイグルマッチング法によって暦年代決定を行った。
測定されたC14年代は較正曲線を用いて暦年代に変換される。較正曲線の変動はその凸凹した形状からwiggleと呼ばれる(図2)。このwiggleのため、C14 年代の年代較正を行った時、得られる暦年代が複数候補あったり、誤差がC14年代のものと比べ非常に大きくなったりする場合がある。しかし、1試料内で年代間隔の分かった複数の試料を得ることが可能な樹木の場合、逆にこのwiggleを利用することによって高精度・高正度の年代較正を行うことができる。(中村 2003)
3−4 ウイグルマッチング試料の採取法
ウイグルマッチング用試料は年輪幅単位でサンプリングを行った。
サンプルの木口面から丸ノコを使って傾斜状に切れ目を2本入れ、正三角柱状にサンプルを切り出し、切り出したサンプルを年輪計測した。その後、計測結果を参照し読み間違いが無いように三角柱状サンプルの頂点部分からカミソリを使って年輪界に沿って1年輪幅(年輪幅が小さく不可能な場合は5年輪幅)ごとに1辺1cmほどの正三角形状のサンプルを切り出した(図3)。年代測定用試料の選出は、ウイグルマッチングに使用する較正曲線(図2)を見てサンプル間の年代幅を考慮し、選出した。C14年代測定は加速器分析研究所で加速器質量分析計(AMS)にて行われた。
3−5 ウイグルマッチングの方法
測定されたC14年代の1年毎における暦年代確率分布をINTCAL04 data set(Stuiver et al,2004)及びCalb Rev5.0(Stuiver and Reimer, 1993)を用いて確率分布Pを計算した。この時iを試料番号、jを暦年代とするとiの暦年代がjである確率分はPi(j)で表せる。この時年輪からそれぞれの試料の相対年代は分かっているため最外年輪の暦年代nが決定されると試料の暦年代も決定される。よって試料番号が1〜5、最外年輪から数えた各試料の年数をそれぞれA〜E年としたとき、最外年輪の暦年代の確率分布Q(n)は、次式に示される確率の積によって定義される。
Q(n)=P1(n−A)×P2(n−B)×P3(n−C)×P4(n−D)×P5(n−E)
ここでnは木柱の最外年輪の歴年代とし、i=1は最外年輪から数えてA年目であり、最外年輪の歴年代がnの時i=1の年輪の歴年代がnーAである確率はP1(n−A) で与えられる。最外年輪の暦年代であるnの範囲を変えてこの積Q(n)が最大になる歴年代nを求める。その後Q(n)の総和を1に規格化し、最大値から確率の和が0.95になる暦年代の範囲を求める。これが95%の確率で示される可能性の高い歴年代である。
4. 結果
4−1 年輪計測結果
4−1−1 自然木の年輪計測結果
自然木の年輪計測データを表1にまとめた。樹種はスギが58点と一番多く、この結果は花粉分析の結果と一致する。次にトネリコが多く29点、次にモクレン属、コナラ属アカガシ亜属がつづく。年輪数はスギが平均80年を超え、年数も最大252年と多かった。湿生の林であったと考えられるが、カエデやクリなどの樹種も含まれる。樹皮に関してはほとんどの個体で失われていたため、死亡年を詳細に議論することは出来ない。クリの年数は少ないものの他の樹種に比べ平均年輪幅が広く、成長が良い。他の樹種は最大で100年前後であった。なお、コナラ属アカガシ亜属に関しては全てのサンプルで部分的に年臨界が不鮮明で、計測が不可能であった。なお巻末に計測個体のリストを付加した。
4−2 相対年代関係
4−1−2 木柱の年輪計測結果
結果を表2に示す。樹種は全てスギである。木柱の場合最外部の年輪は伐採年に相当するため重要となるが、8個体中1個体のみしか確認できなかった。そのため伐採年を詳細に議論することは出来ない。年輪数も平均53年と少ないが、スギ自然木と比較してみると平均年輪幅が2.52で倍以上直径も年数に対して大きく、成長が早いことが分かった。
4−2−1 自然木の相対年代関係
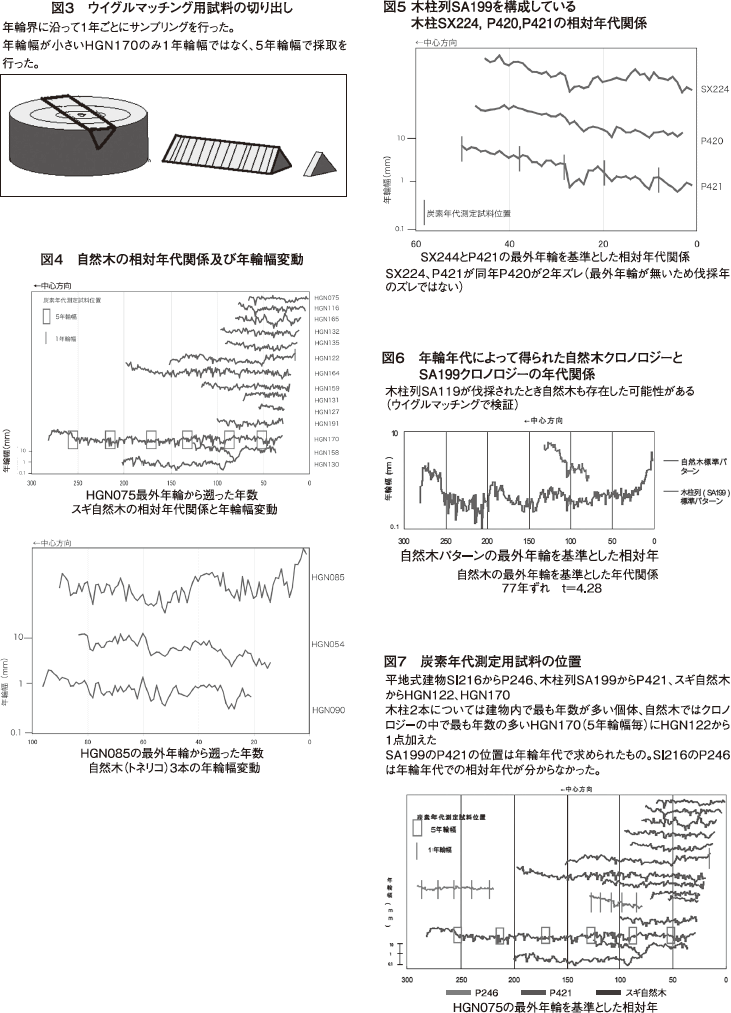
自然木の相対年代関係結果を図4に示す。自然木では同じ樹種内の個体間でクロスデーティングを行い、個体内での相関が得られた個体同士をクロスデートし、t値が3.5以上の組み合わせを組んでいった。その後組み合わせ同士でのクロスデーティングを行い、各個体間の年代関係を明らかにしていった。こうして相対的な年代関係が明らかになった個体の年輪変動のパターンを標準化し、クロノロジーを作成した。その結果、自然木に関してはスギとトネリコのクロノロジーを組むことが出来た。
自然木クロノロジーでのt値を表3に示す。HGN164とHGN 170がt値10.94と最も高くなった。HGN 164は年数の少ないHGN127以外の個体とt値4〜6で高い相関を示し、その他の個体も複数個体と4以上の値を示している。スギでは計測を行った58個体中比較的年数の多い14個体での281年分のクロノロジーとなった。トネリコでは計測した29個体中3個体のみで100年を超えた個体を含んだクロノロジーを組むことが出来なかった。
この結果からスギとトネリコの自然木クロノロジーを平均・標準し、マスタークロノロジーを作った。この際スギHGN127の年数が少なく2個体以外の相関が悪く(表3)、不確かなため除きスギでは13個体でマスタークロノロジーを作成した。しかし、マスタークロノロジーは281年分のうち約80年分がHGN170 1本のみで構成されている。
モクレン属では木口面画像では年臨界が読み取れず、切片を作成した結果年輪を読み取ることが出来た。計測を行った個体のうち2個体に相関が見られた。このことからモクレン属の切片採取による年輪解析は有効であることが分かった。
4−2−2 木柱の相対年代関係
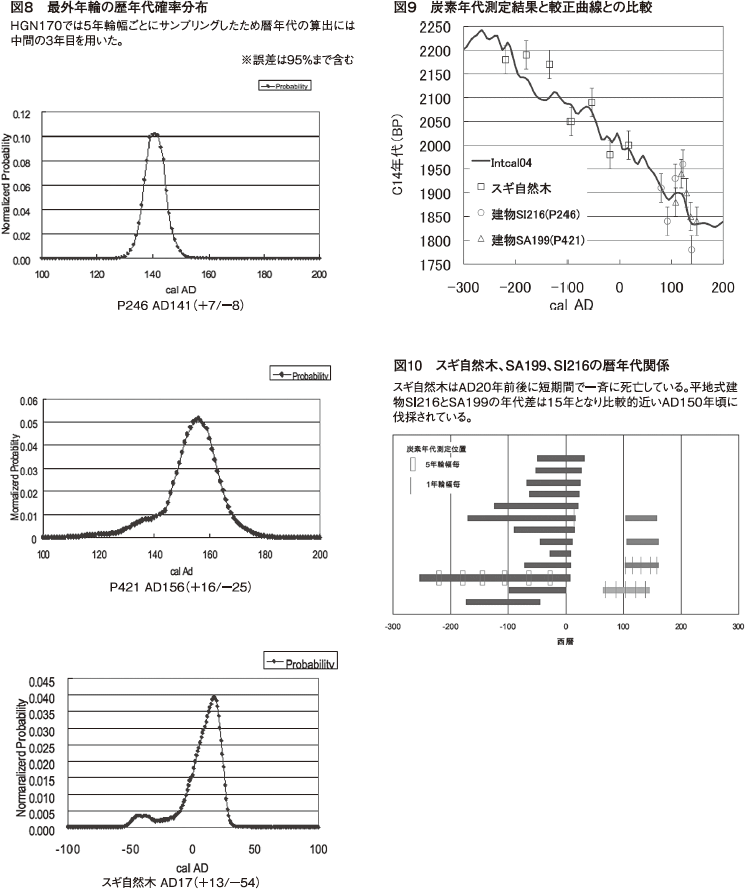
木柱列SA199内木柱間のクロスデーティング結果を図5に、t値を表4に示す。P421とSX224が同一年、P420が2年ずれとなった。3個体とも最外年輪は失われているため、伐採年は分からないが、ほぼ同時期に伐採されたものと考えられる。これは同一建物内であることから信頼性のある結果といえる。この結果を元にSA199のクロノロジーを作成した。
平地式建物SI216では木柱4本内での高い相関は得られず、また、掘立柱建物SB341は1本のみである。次に、異なる建物同士でのクロスデーティングを行ったが、各建物間での高い相関は得られなかった。主な原因としては木柱の年輪幅変動が変化に乏しかったことと、年数が少ないためと考えられる。なお偽年輪、不連続年輪等は見られない。
4−3 自然木と木柱との相対年代
SA199とスギ自然木のクロノロジー同士のクロスデーティング結果を図6に示す。ずれは77年でt値は4.28を示した。これは木柱列SA199の木柱が生育、伐採時に自然木が存在していた可能性を示している。この結果を検証するためウイグルマッチングによる暦年代決定を行った。
4−4 ウイグルマッチング結果
炭素年代測定試料位置を図7に示す。試料は平地式建物SI216からP246、木柱列SA199からP421とスギ自然木からHGN122、HGN170の計4本である。木柱2本については建物内で最も年数が多い個体、自然木ではクロノロジーの中で最も年数の多いHGN170を計測対象とした。炭素年代測定用サンプルはHGN170のみ5年輪幅ずつ採取した。
C14年代測定結果を表5に示す。試料採取位置の番号が測定サンプルの最外年輪を1とした時の年数に当たる。
最外年輪の歴年代nを変化させたときの確率の積Qの分布を図8に、較正曲線(Intcal04)と測定結果との比較を図9に示す。最外年輪の暦年代はP246でAD141 (+7/−8)、P421でAD156(+16/−25)、スギ自然木がAD17(+13/−54)となった。誤差は95%の確率まで含めたものである。
5. 考察
5−1 出土木材の暦年代決定
木柱列SA199 、平地式建物SI216 とスギ自然木との暦年代結果を図10に示す。木柱列SA199とスギの自然木との年輪年代結果では自然木と年代が重複している可能性があり、ウイグルマッチングで検証したが、ウイグルマッチングの結果では木柱の最外年輪がAD141( +7/−8)、自然木がAD17(+13/−54)となり自然木と木柱では年代が重複していない結果となった。クロスデーティングではt値は4.28と、相関が見られていたがウイグルマッチングの結果を見ると自然木は木柱が生育をはじめた時代にはすでに死亡していた。また、木柱の最外年輪の暦年代はSA199の木柱P421がAD156( +16/−25)、SI216の木柱P246がAD141(+7/−8)とどちらも弥生時代後期頃の値を示した。共に伐採された年の年輪が失われているため伐採年を断定することは出来ないが、この結果をみるとこれらの木柱がAD150年前後の短い期間に伐採された可能性が極めて高いと考えられる。
5−2 木柱と自然木との年代関係
スギ自然木はAD20年付近で多くの個体が短期間に死亡しているが、多くの個体は樹皮が無く死亡年の詳細な判断は出来ない。しかし数十年以上失われている個体は無いため、短期間で死亡した年代の誤差は少ない。木柱の年代は伐採年がAD150年前後であることから自然木が死亡した後に生育したと考えられる。これらの結果は遺跡で行われた花粉分析での、III層下部ではスギ優勢であるがコナラ属アカガシ亜属など照葉樹があり、III層上部では水田の拡大とともにスギ林の減少が見られるという結果とも一致している。
年輪計測から自然木と木柱の成長速度に大きな差があり、スギ自然木が平均年輪幅1.05mmに対し、木柱は2.52mmと倍以上あり、木柱は自然木に比べ非常に成長が速い。自然木に比べ光環境の良い場所で木柱が生育したことが分かった。このことから、埋没林はスギ優占でコナラ属などを伴う林であったが、何らかのイベントによってAD20年頃短期間に枯死、埋没した。その後AD150年頃埋没林周囲に人が住み始め、周囲に水田が広がっていったことが推測できる。
しかし、計測を行ったスギ自然木の年輪計測で、個体内での相関がよく正しく計測できたと考えられる個体は58個体中52個体であるのに対して、相対関係を求められたのは14個体と少ない。年数の少ない個体で高い相関が得られなかったと考えられるが、100年を超えた個体全9個体中、相対関係を求められたのは4個体と半分程度であった。同時期に死亡した個体であればこうした年数の多い個体で相関が見られないことは考えにくいが、14個体以外での相関の高かった組み合わせも無かった。これらから偽年輪や、不連続年輪などによって年輪の読み飛ばし等により、相関が悪くなっている可能性が考えられる。相関が得られた個体内でも偽年輪や不連続年輪があり、クロノロジーを組んでゆく過程で発見したものもあった。相関が求められなかった個体では実際には同年代に存在していたが、こうした偽年輪、不連続年輪が多く高い相関を求めることが出来なかった可能性が大きい。
5−3 建物間の年代関係
SA199とSI216は年輪年代で相対年代を求めることが出来なかったが、ウイグルマッチングによって15年差という結果が求められた。このため年輪年代でこの年代差での相関の再確認を行った。しかし、どの木柱間の組み合わせでも、高い相関は見られなかった。
5−4 クロノロジーの構築について
古墳時代前期のスギにおける280年以上のクロノロジーを組むことが出来た。このクロノロジーの構築は今後新たに発見された古墳時代の遺跡でのクロノロジーと繋がる可能性や、出土木材の年輪年代決定に適用できる可能性がある
引用・参考物件
新潟県埋蔵文化財調査事業団(2008) 姫御前遺跡I 北陸新幹線関係発掘調査報告書VIII.
新潟県埋蔵文化財調査事業団(2009) 自然科学分析業務報告書. 4-5
加速器分析研究所(2009) 姫御前遺跡IIにおける放射性炭素年代(AMS測定)
木村勝彦、斉藤智治、中村俊夫(2004) 青田遺跡における木柱の年輪年代解析による建物群の年代関係の検討.
中村俊夫、石塚友希夫、奥野充、木村勝彦、金伯禄、森脇広(2003) 白頭山火山の10世紀における巨大噴火の高精度AMS14C年代測定 名古屋大学加速器質量解析計業積報告書(XIV).
中村俊夫(2002)高精度14C年代測定と14C年代の歴年代較正 第1回高精度14C年代測定研究委員会 公開シンポジウム: 2−7.
木村勝彦(2002)縄文時代の高精度編年に向けて 年輪年代学からの取り組み 第1回高精度14C年代測定研究委員会 公開シンポジウム: 19−22.
Schweingruber,F.H.(1989) Tree Ring :49-51.
Baillie M.G.L.(1982) Tree-Ring Dating and Archaeology. The University of Chicago
ポーラ・ルダル(1992)(鈴木三男・中田裕美訳、1997)植物解剖学入門.
島地 謙 伊藤隆夫(1982) 図説 木材組織:22−27 42−47 72−73.
澤田恵美(2005)14Cウイグルマッチング法を用いた白頭山噴火の高精度での年代決定. 福島大学教育学部学校教育教員養成課程 卒業論文

|
